Gather and Sow: June 2026
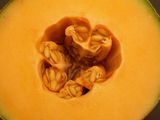

Welcome to June, when strawberry season is finally well underway across wide swaths of the Northern Hemisphere. With so many strawberries in so many kitchens this month, now is the perfect time to merge botany lab and breakfast preparation by working through the many parts of a strawberry. We kick off the strawberry love with some explanations in Food for Thought. You can learn quite a bit about flowers by cutting up a strawberry, as we do in this month’s Botany Lab of the Month. As it turns out, this enigmatic little gem is packed with coincidences and apocrypha along with its citric acid and anthocyanins. We conclude with recent relevant media and journal articles in Gleanings and Baker’s Dozen.
Strawberries are in the rose family (Rosaceae), along with numerous other fruity stars of summer and early fall: the stone fruits (cherries, peaches, plums, almonds); the brambles (raspberries, blackberries, and their numerous wild relatives); the pomes (apples, pears, quince); and several other delightful species (loquat, medlar, serviceberry, rosehips, rowan, hawthorn). We will continue to explore the rose family in this summer’s Gather and Sow newsletters.
Food for Thought
Coincidences and confusions about strawberries
Confusions
You may have heard that strawberries are not berries, but what is a berry? Colloquially, a berry is a smallish, sweet, juicy thing plucked off a plant. Strawberries comfortably fit that description. The problem arises from the much narrower definition of berry employed by botanists (with further refinement and debate among the most pedantic ones) (1, 2). A berry-type fruit is fleshy throughout, without any hard or papery bits, such as the pit of a peach or the papery insides of an apple. Grapes are berries, as are persimmons. Other familiar berries include eggplants, avocados, and tomatoes. Strawberries are not built this way, as we demonstrate below in Botany Lab of the Month: The botanists’ way to cut up a strawberry.
But strawberry confusion, both botanical and historical, goes way beyond the falseness of the fruit. For example, the modern horticultural strawberry is called Fragaria x ananassa, the “pineapple strawberry.” The “x” in the name marks its hybrid origin. Early strawberry breeders apparently thought that the hybrid shared some flavors with pineapples (3, 4). Interestingly, pineapples – which are obviously neither pines nor apples – are true berries. The large spiny yellow-green structure is an aggregate of berries, nestled down into a flowering axis.
Although both parent species of our current commercial strawberry came from the Americas – the previously domesticated F. chiloensis from Chile and wild F. virginiana from the eastern U.S. – the first hybridization between them happened in France in the 17th century (5–13). This may be why we so often associate strawberries with France. The original French strawberry, the fraise de bois, was very popular until it was replaced first by F. virginiana and then by the hybrid pineapple strawberry.
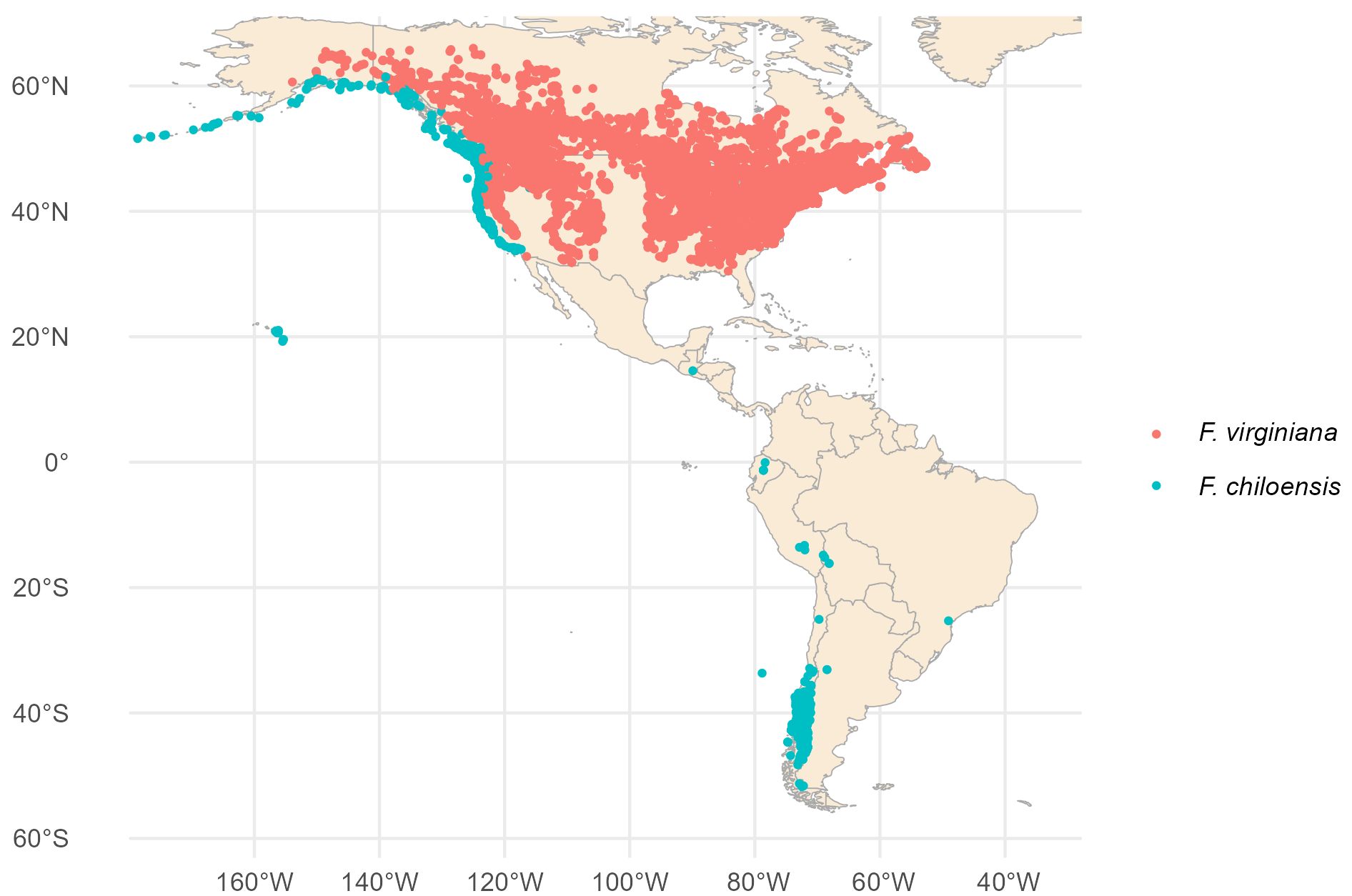
Coincidences?
It was an 18th century French spy sent to Chile and Peru by King Louis XIV who was responsible for bringing the Chilean species to France where it could cross with the Virginia species (7). This spy’s name was Amédée Frézier. Frézier looks and sounds remarkably like “fraisier,” the French word for a strawberry plant (and a strawberry cake). Frézier is a variant of Fraser (Frasier, Frazier, Frizzelle, etc.), a Scottish clan with its roots in France, whose heraldry features strawberry flowers.
It is interesting enough that Captain Frézier was predestined by his family name to be the father of the modern strawberry, but it might be even more so. One story holds that in the early 10th century, a French nobleman hosted a feast for King Charles III of France and an Italian Cardinal, who were traveling together. The nobleman ended the meal with a simple plate of perfect strawberries, which so impressed the king that he gave the nobleman the name Frasier and a coat of arms bedecked with strawberry blossoms.
And what was the nobleman’s original name? Julius de Berry.
For further reading, Atlas Obscura has a fun article about Frézier and Antoine Duchesne, the botanist at Versailles who realized that the two American species in the palace garden had hybridized and made excellent fruit. They also have an article about frutilla blanca, the large, white domesticated form of F. chiloensis that Frézier took back to France.

Botany Lab of the Month
The botanists’ way to cut up a strawberry
For this exercise, all you really need is a fresh strawberry (the fresher the better) and a small paring knife. Reading glasses or a hand lens might be helpful, too.
1. Examine the whole berry. The first thing to notice about a whole strawberry is its relationship to the flower it came from. The stem where it was attached to the plant was originally the stem holding up the flower; so if the stem is pointing up, you have to imagine a flower pointing downwards. Or you can turn the berry stem-side down.

2. Look at the green fringe of sepals. As a child, Katherine called this green whorl the “Kermit collar,” in honor of Kermit the Frog, who also appears to wear a ring of sepals. Sepals are the outermost ring of flower parts, and they often serve to protect the flower when it is still a bud. Collectively, the sepals are called a calyx.
Although it looks like there are ten sepals in two alternating rings of five, there are not. The outer ring is called an epicalyx and is actually composed of stipules, appendages of the sepals. And there’s even more deception than that. Each sepal is flanked by two stipules, and the adjacent stipules from neighboring sepals fuse together, giving the impression that single stipules alternate with the sepals. But fusion during development is kind of sloppy and can be incomplete, sometimes leaving the total number of green appendages a bit more or less than ten. (The whole rose family is characterized by stipules, which are easily seen as long flaps of tissue at the base of a rose leaf).

3. Lift the sepals (calyx) to reveal the stamens. The whorl of appendages just inside the sepals was composed of petals, but these are gone by the time strawberries have matured. Occasionally, though, a shrunken and mildewed white petal is still stuck on the strawberry somewhere. What you will always be able to see under the sepals is a couple of rings of stamens, which remain firmly attached to the calyx. Strawberry stamens are paddle-shaped, the wide part being the anthers, where pollen is produced. By now, the anthers will have opened up to shed their pollen, and those slits should be visible along the edges of the anthers.
4. Look very closely at the surface of the “berry” and question the way we normally talk about it. Here’s the “critical thinking” part of the exercise. Just inside the ring of stamens we expect to see one or more pistils, whose most important part is the ovary. Ovaries are the structures that contain the newly-fertilized eggs on their way to becoming seeds. The ovaries themselves are triggered by the seeds to mature into fruits.
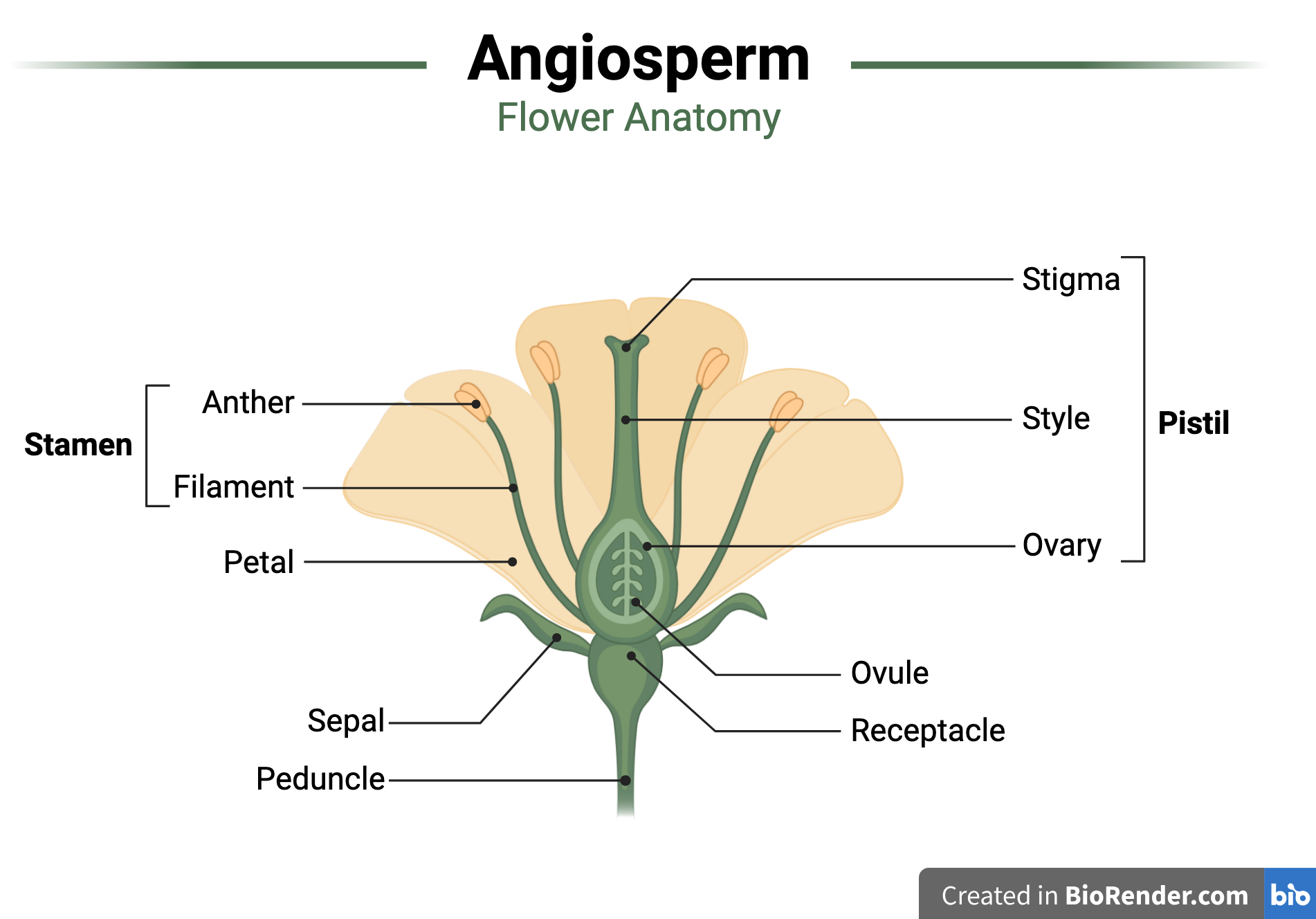
If we follow the familiar botanical blueprint, we might start by guessing that the big red mound in the center of the strawberry flower has to be the mature ovary (the fruit). But it is covered in tiny tan hard bits resembling seeds. And we know perfectly well that seeds do not grow on the outside of a fruit. Making seeds on the outside of an ovary seems like a very big violation of expected developmental rules, even bigger than producing an unexplained mound where we have gone looking for an ovary.

However, if you look very closely at one of those “seeds,” you will see that it is tipped with a tiny pin-headed thread. That thread is a style leading to an ovary. Any pollen grain landing on the stigma – the pin-head top – could germinate and develop a tube to digest its way through the style to deliver a sperm cell to the egg cell in the ovary.
The presence of a stigma and style tells us that each of those “seeds” is actually an individual matured ovary, and the strawberry mound is thus covered in many many separate tiny hard fruits. We call those fruits achenes (ay-keens) so that we can make jokes about our achene hearts (and because they are a type of single-seeded indehiscent fruit derived from a superior ovary). Although most generic diagrams of the typical flower include only a single ovary, in the real world flowers can include several ovaries from several separate pistils. That’s what strawberry flowers do.

5. Finally, put the knife into the “berry.” Now that we know that the red mound is not the fruit, and that the real fruits are called achenes and not berries, we know that we are definitely getting at least 5 fruits a day when we eat strawberries. What we get from the not-really-a-fruit red mound, though, is the sweet, tart, juicy, vitamin-packed object of our spring dreams. Botanically, the red mound is a very thick flower axis called a receptacle, upon which the sepals, petals, stamens, and pistils develop.
Slice the strawberry lengthwise and notice the white lines running out to the surface of the receptacle where they feed directly into the achenes. Those are veins (and their associated tissues), feeding the developing seeds inside the achenes. The hollow center results when the outer part of the receptacle grows faster than the inner part and tears away from it.

Strawberries vs. blackberries and raspberriesTo summarize, then, a strawberry (not a berry) is a fleshy sweet floral axis (receptacle) covered in many separate dry fruits called achenes. It is conceptually helpful to contrast strawberries with the closely related caneberries (or brambles), which include blackberries and raspberries (Rubus species). The caneberry brambles will be the topic of next month’s newsletter, so consider this anatomy lesson a preview. Like strawberries, caneberries have an enlarged receptacle covered in individual fruits; however, unlike strawberries, caneberries have a dry, bland receptacle. In caneberries, it is the actual fruits that are fleshy and sweet. Although the fruits are not achenes, they are also not properly called berries. Caneberry fruits have a fleshy mesocarp (“middle fruit”) surrounding a pit (a stony endocarp), containing a single seed. They are essentially miniature plums. We will cover this type of fruit – a drupe – when we talk about stone fruits, later this summer. Raspberries seem hollow because their receptacles stay behind on the bush, and the little drupes cohere in a beautiful jeweltone dome. Blackberry receptacles are harvested and eaten with the rest of the fruits. And to get even more technical, because the fruits are adorably small and collected together in a single flower, they are more accurately called an aggregate of drupelets.
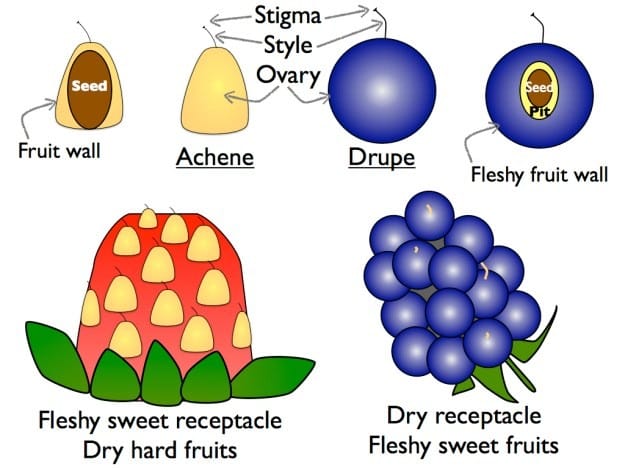
Strawberries and caneberries use different strategies to the same end, but both are effective. Whether it is the receptacle of the strawberry or the drupelets of the caneberry, a sweet and fleshy structure attracts and rewards seed dispersers while a small hard covering – the achene wall or the drupe pit – protects the actual seeds against being digested.
Gleanings and Baker’s Dozen
Gleanings
- They really do consider everything, even the importance of plant polyploidy! A cool new paper featured by National Public Radio (or Nifty Polyploid Radio)
- Good journalism on the plight of the Florida citrus industry in Slate
- Gastropod podcast episode with a vignette featuring beets: Feel the Beet: The Most Fascinating Woman You've Never Heard Of
- The Guardian reporting on wheat farming troubles in the US.
- Bolivian farmers facing hardship as quinoa popularity wanes.
- The Edge Effects podcast and website is great. Here is a recent episode about the history of potatoes in Eastern Europe.
- Why Doesn’t Coffee Taste Like Caffeine? from Nautilus. Other molecules in roasted coffee form a complex with caffeine that is too big to interact with bitter taste receptors on the tongue.
- Flowers on the EDGE: the first global assessment of the most unique flowering plants A needed and compelling analysis of angiosperm uniqueness, rarity, and risk. They also did a gymnosperm version. By Kew.
- Books of interest:
- The Hidden Life of Trees: A Graphical Adaptation by Peter Wohlleben and Fred Bernard, and illustrated by Benjamin Flao, based on Wohlleben’s The Hidden Life of Trees
- What the Flora by Vanessa Fuchs, from the Botanic Gardens of Sydney. There’s also a YouTube version.
- Atlas of Botany, published by DK. The map of the spread of Lamiaceae species around the world alone makes this book worth it.
- Water in the Desert, by Gary Paul Nabhan
- The Story of Birds by Steve Brusatte
- Humans: The Evolution of a Species, published by DK, edited by Dr. Alice Roberts
- Forest of the Sea: The Remarkable Life and Imperiled Future of Kelp, by David Helvarg
Baker’s Dozen
- Dominguez Mendez, L., & Studer, A. (2026) Is more better? Polyploidy in crops with diverse end uses and the potential for future applications. Annals of Botany 137: 631-639. As we described above, the hybrid garden strawberry is octoploid, meaning it is a polyploid crop with eight sets of chromosomes. The genetics of polyploid crops has hampered breeding efforts, but this paper explores the potential advantages of polyploidy, especially for improving yield.
- Leung, A., et al. (2026) Convergence, stability, and thermal adaptation of the rubisco large subunit in plants. Evolution. Preprint. Rubisco does not make cookies, it just makes the sugar that goes into the cookies. Rubisco (otherwise known as d-ribulose 1,5-bisphosphate carboxylase/oxygenase) is the enzyme used to fix atmospheric carbon into sugars through photosynthesis. It is said to be the most abundant protein on earth. This paper sheds some light on how this critically important enzyme has evolved to work optimally under different temperature conditions.
- Sun, Y., et al. (2026) Gibberellin-flavonoid crosstalk intensifies symbiotic nitrogen fixation in pea during maize–pea interspecific root interactions. New Phytologist. Preprint. When legumes (such as peas) and maize are grown together, maize benefits from the nitrogen fixed by the roots of the legume. This paper shows that signals from maize roots trigger pea plants to up their N-fixation game by making more nodules where symbiotic bacteria live (and do the actual biochemical work).
- Botero-Delgadillo, E., et al. (2026) Forest cover drives bird occupancy across guilds and habitats in a Colombian coffee-farming landscape. J. Applied Ecology 63: e70414. Growing coffee in the shade, under a native forest canopy, helps conserve animal biodiversity. This paper reports that the diversity and density of the surrounding forest is also very important for supporting bird diversity.
- Wang, T., et al. (2026). StLAX5 regulates stolon initiation to control the tuber number and productivity of potato. New Phytologist online early. Potato tubers grow on thin underground stems called stolons. Plant hormones control the number of stolons and tubers.
- Varotto, S., et al. (2026) From chill to bloom: seasonal coordination of reproductive development in peach and temperate fruit trees. J. Experimental Botany 77: 2835–2850. A lovely review of the transition from winter to spring in temperate fruit trees.
- Sazon, L. A. R., et al. (2026) Stem nitrogen accumulation through vegetative storage proteins and mobilization to seeds supports high-yielding soybean. J. Experimental Botany 77: 2998–3008. We know that legumes, and soybeans in particular, are a hugely important source of protein. Proteins have a lot of nitrogen in them. Soybean plants have to store nitrogen throughout their growth and flowering phase in order to have enough to pack their seeds with protein. It turns out they are storing a lot of nitrogen in their stems.
- The latest special issue of the Journal of Experimental Botany is all about wheat. The article by Che et al. on seed storage proteins (SSP) has a wonderful graphic:
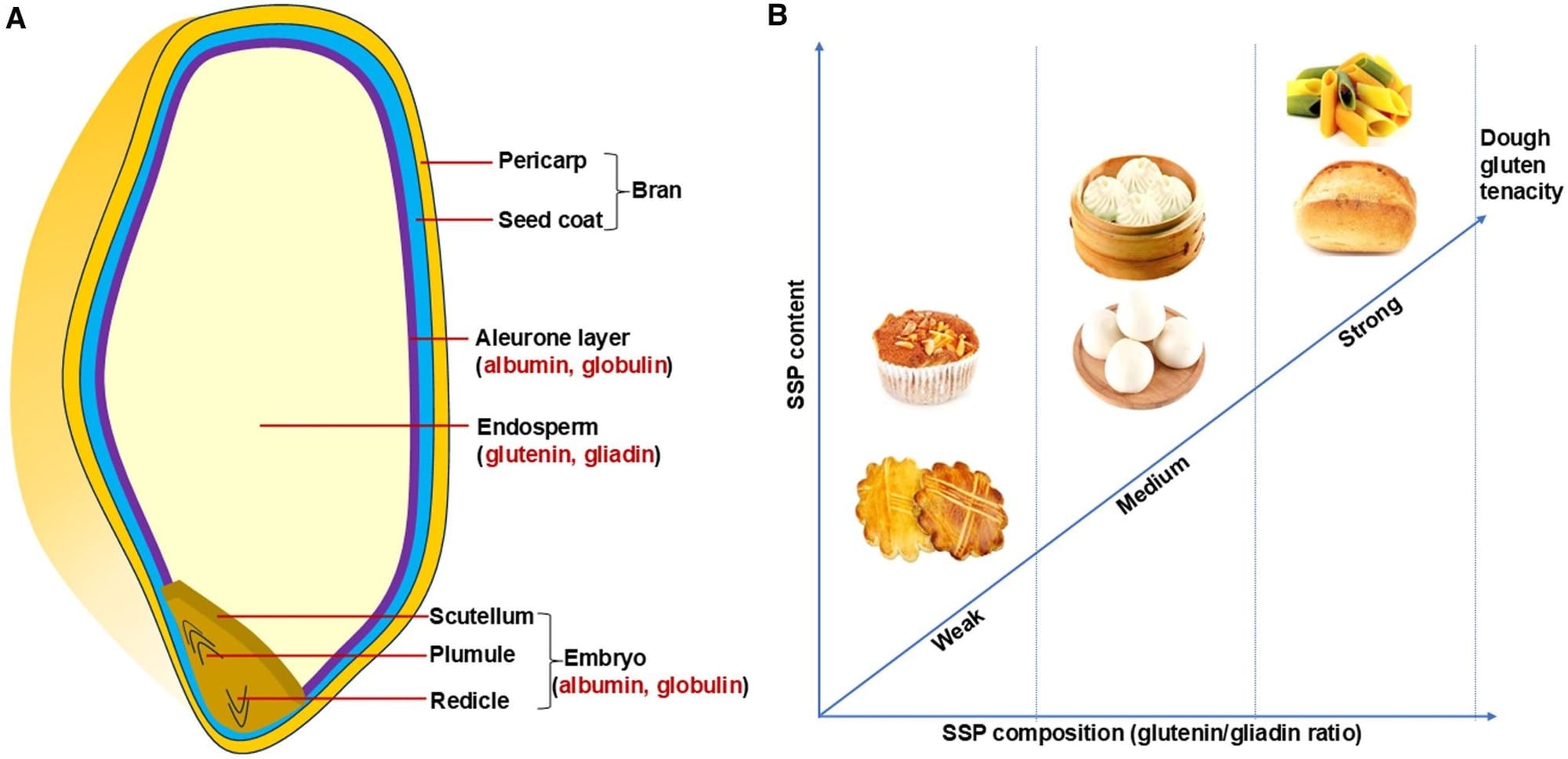
- PNAS has a special feature about polyploidy and stress with several articles about crop plants.
- Rusishvili, N., et al. (2026) An independent center for the origin of bread wheat in the Neolithic period of Georgia in the South Caucasus. PNAS 123: e2537697123. Archaeological evidence that wheat domestication in the South Caucuses coincided with grape domestication. Bread and wine! So cheers the commentary on the article.
- Lu, Y., et al. (2026) Soil temperature fluctuations modulated millet agriculture evolution in Neolithic East Asia. PNAS 123: e2529151123. Soil temperature, not air temperature or precipitation, may have been determinative about early patterns of agriculture in East Asia.
- There are two recent papers about neighbor detection and plant responses in crops. Crop plants are typically sown at high densities, even though the resulting competition between neighboring plants for light and soil resources may negatively affect yield. Plants may alter their growth and development in responses to neighbor detection. Light quality is one of the environmental cues plants use to detect neighbors. Phytochromes are molecules that plants use to perceive light quality. Their action is hugely significant for stand development in natural and agricultural systems. The ability of plants to change their morphology in response to their environment is called plasticity. Phytochrome-mediated plasticity first came on our radar through the work of Johanna Schmitt. One of her foundational papers is our choice for this month’s Oldie but Goodie, below.
- Surya, K., et al. (2026). Phytochrome-mediated shade-avoidance responses and its impact on growth and physiology in crops—A review. Photochemistry and Photobiology. Pre-print.
- Pereyra, M. E., et al. (2026). Avoiding the shadow: how plants perceive neighbors and reshape the crop light environment. Plant Physiology 200: kiag034.
- Oldie but Goodie: Schmitt, J., et al. (2003). The Adaptive Evolution of Plasticity: Phytochrome-Mediated Shade Avoidance Responses. Integrative and Comparative Biology 43: 459-469. Plants may change their growth and development in response to environmental perception, a phenomenon called plasticity. As we mentioned above, plants use phytochromes to perceive light quality, one of the environmental cues that affects plant growth. Phytochrome-mediated changes in plant growth and development are examples of plasticity. The quality of light changes when it passes through a leaf. Phytochromes can detect this difference, allowing a plant to determine whether or not it is located underneath or near the canopy of another plant. If a plant is shaded by another plant, phytochrome mediates a whole cascade of shade avoidance responses, which usually involves either hunkering down or putting all energy into height growth to race to the top of the canopy. Shmitt’s key insight is that this flexibility in growth, this plasticity, is itself adaptive.
References
1. Spjut, Richard W., A Systematic Treatment of Fruit Types (New York Botanical Garden, New York, USA, 1994; http://www.worldbotanical.com/fruit_types.htm#What%20is%20a%20Nut).
2. A. V. F. Ch. Bobrov, M. S. Romanov, Morphogenesis of fruits and types of fruit of angiosperms. Bot. Lett. 166, 366–399 (2019).
3. J. Yan, Z. Ban, H. Lu, D. Li, E. Poverenov, Z. Luo, L. Li, The aroma volatile repertoire in strawberry fruit: a review. J. Sci. Food Agric. 98, 4395–4402 (2018).
4. M. Porter, Z. Fan, S. Lee, V. M. Whitaker, Strawberry breeding for improved flavor. Crop Sci. 63, 1949–1963 (2023).
5. J. Janick, “The Origins of Fruits, Fruit Growing, and Fruit Breeding” in Plant Breeding Reviews (John Wiley & Sons, Ltd, 2005; https://onlinelibrary.wiley.com/doi/abs/10.1002/9780470650301.ch8), pp. 255–321.
6. M. A. Hardigan, A. Lorant, D. D. A. Pincot, M. J. Feldmann, R. A. Famula, C. B. Acharya, S. Lee, S. Verma, V. M. Whitaker, N. Bassil, J. Zurn, G. S. Cole, K. Bird, P. P. Edger, S. J. Knapp, Unraveling the Complex Hybrid Ancestry and Domestication History of Cultivated Strawberry. Mol. Biol. Evol. 38, 2285–2305 (2021).
7. Darrow, G. M., The Strawberry: History, Breeding, and Physiology (Holt, Rinehart & Winston, New York, USA).
8. C. E. Finn, J. B. Retamales, G. A. Lobos, J. F. Hancock, The Chilean Strawberry (Fragaria chiloensis): Over 1000 Years of Domestication. doi: 10.21273/HORTSCI.48.4.418 (2013).
9. P. P. Edger, T. J. Poorten, R. VanBuren, M. A. Hardigan, M. Colle, M. R. McKain, R. D. Smith, S. J. Teresi, A. D. L. Nelson, C. M. Wai, E. I. Alger, K. A. Bird, A. E. Yocca, N. Pumplin, S. Ou, G. Ben-Zvi, A. Brodt, K. Baruch, T. Swale, L. Shiue, C. B. Acharya, G. S. Cole, J. P. Mower, K. L. Childs, N. Jiang, E. Lyons, M. Freeling, J. R. Puzey, S. J. Knapp, Origin and evolution of the octoploid strawberry genome. Nat. Genet. 51, 541–547 (2019).
10. A. Liston, R. Cronn, T.-L. Ashman, Fragaria: A genus with deep historical roots and ripe for evolutionary and ecological insights. Am. J. Bot. 101, 1686–1699 (2014).
11. K. E. Hummer, N. Bassil, W. Njuguna, “Fragaria” in Wild Crop Relatives: Genomic and Breeding Resources: Temperate Fruits, C. Kole, Ed. (Springer, Berlin, Heidelberg, 2011; https://doi.org/10.1007/978-3-642-16057-8_2), pp. 17–44.
12. Q. Qiao, P. P. Edger, L. Xue, L. Qiong, J. Lu, Y. Zhang, Q. Cao, A. E. Yocca, A. E. Platts, S. J. Knapp, M. Van Montagu, Y. Van de Peer, J. Lei, T. Zhang, Evolutionary history and pan-genome dynamics of strawberry (Fragaria spp.). Proc. Natl. Acad. Sci. 118, e2105431118 (2021).
13. K. A. Bird, M. A. Hardigan, A. P. Ragsdale, S. J. Knapp, R. VanBuren, P. P. Edger, Diversification, spread, and admixture of octoploid strawberry in the Western Hemisphere. Am. J. Bot. 108, 2269–2281 (2021).
14. GBIF, Fragaria occurrence data; https://doi.org/https://doi.org/10.15468/dl.4g9ake.
15. J. F. Hancock, H. H. Prince, Long-distance dispersal of the beach strawberry, Fragaria chiloensis, from North America to Chile and Hawaii. Ann. Bot. 127, 223–229 (2021).
16. Massicotte, P., South, A., rnaturalearth: World Map Data from Natural Earth, version 1.2.0 (2026); https://ropensci.r-universe.dev/rnaturalearth.
17. A. C. Willoughby, Rapid strawberry domestication left room to grow. Plant Cell 36, 1570–1571 (2024).
18. A. Gudowska, A. Cwajna, E. Marjańska, D. Moroń, Pollinators enhance the production of a superior strawberry – A global review and meta-analysis. Agric. Ecosyst. Environ. 362, 108815 (2024).
19. Z. Fan, V. M. Whitaker, Genomic signatures of strawberry domestication and diversification. Plant Cell 36, 1622–1636 (2024).
20. M. A. Hardigan, M. J. Feldmann, A. Lorant, K. A. Bird, R. Famula, C. Acharya, G. Cole, P. P. Edger, S. J. Knapp, Genome Synteny Has Been Conserved Among the Octoploid Progenitors of Cultivated Strawberry Over Millions of Years of Evolution. Front. Plant Sci. 10 (2020).
21. Angiosperm Phylogeny Website. https://www.mobot.org/MOBOT/research/APweb/.



